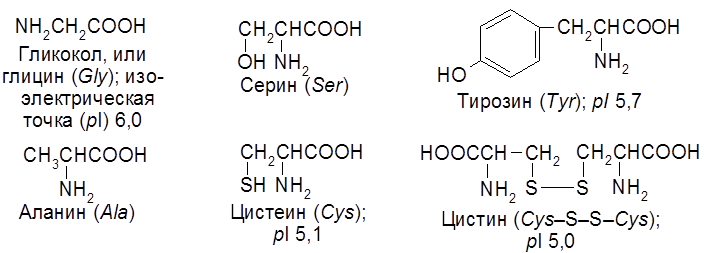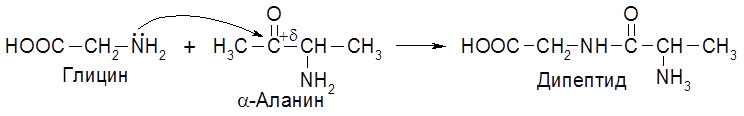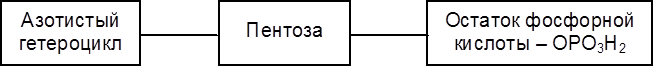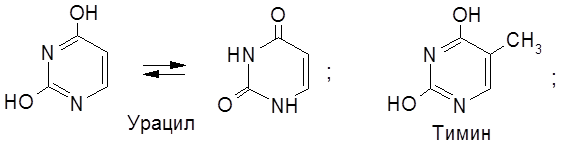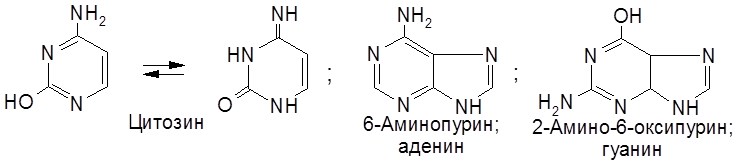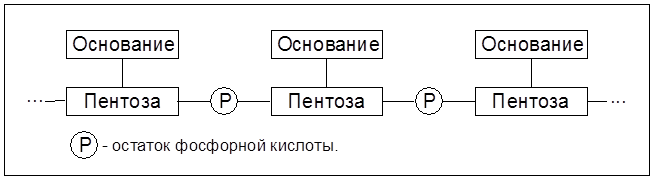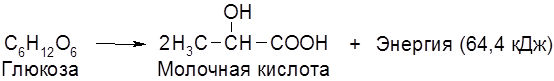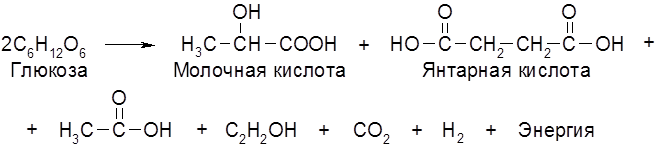| МЕТОДИЧЕСКИЕ УКАЗАНИЯ по изучению биологической химии и задания для самостоятельной работы |
|
| Добавил(а) Administrator | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 16.02.11 13:09 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
МЕТОДИЧЕСКИЕ УКАЗАНИЯ
по изучению биологической химии и задания Для студентов заочного отделения по специальностям: – технология жиров и жирозаменителей – 6.091.700 – технология бродильных производств и виноделия – 6.091.700 – технология хранения и переработки молока – 6.091.700
СОДЕРЖАНИЕ1. ВВЕДЕНИЕ.. Цель и задачи методических указаний. 2. ВИТАМИНЫ... 2.1. Общая характеристика. 2.2. Водорастворимые витамины.. 2.2.1. Витамин С.. 2.2.2. Никотиновая кислота. 2.2.3. Рибофлавин (витамин В2) 2.3. Жирорастворимые витамины.. 2.3.1. Витамин А (ретинол, антиксерофтальмический витамин) 2.3.2. Витамин D (холекальциферол, антирахитичный) 2.3.3. Витамин Е (токоферол, антистерильный витамин). Антиоксиданты.. 2.3.4. Витамин К (филлохинон, антигеморрагический витамин) 2.4. Вопросы для самоконтроля. 3. фЕРМЕНТЫ, КОФЕРМЕНТЫ, ГОРМОНЫ... 3.1. Общая характеристика ферментов. 3.2. Строение ферментов. 3.3. Классификация и номенклатура ферментов. 3.4. Коферменты.. 3.5. Гормоны, фитогормоны.. 3.6. Контрольные вопросы.. 4. Биоэнергетика. Биологическое окисление.. 4.1. Общая стратегия энергообеспечения организмов. 4.2. Переносчики химической энергии и некоторые 4.3. Анаэробный и аэробный механизм окисления – 4.4. Цитохромы и их предшественники. Другие конечные оксидазы.. 4.5. “Прямые” оксидазы.. 4.7. Вопросы для самоконтроля. 5. Углеводы – строение, свойства, биологическая роль.. 5.1. Наиболее важные представители – строение и свойства. 5.1.1. Моносахариды.. 5.1.2. Олигосахариды. Дисахариды.. 5.1.3. Полисахариды.. 5.2. Обмен углеводов – анаболизм и катаболизм. 5.2.1. Гликолиз. 5.2.2. Пировиноградная кислота. 5.2.3. Цикл трикарбоновых кислот (цикл Кребса) 5.2.4. Глиоксилатный цикл. 5.2.5. Пентозофосфатное (апотомическое) окисление глюкозы.. 5.2.6. Особенности обмена углеводов в организмах 5.2.7. Анаэробный распад (брожение) углеводов (сахаров) 37 5.2.8. Фотосинтез. 5.2.9. Хемосинтез. 5.3. Вопросы для самоконтроля. 41 6. Липиды – химическое строение, свойства и биологическая роль. кАТАБОЛИЗМ И АНАБОЛИЗМ... 42 6.1. Нейтральные жиры.. 6.2. Фосфолипиды (фосфоглицериды) 6.3. Сфинголипиды.. 6.4. Стериды и стерины.. 6.6. Обмен липидов. 6.7. Вопросы для самоконтроля. 7. Белки и аминокислоты – строение, 7.1. Общая характеристика. Биологическая роль. 7.2. Аминокислоты – строение и свойства. 7.3. Структура и классификация белков. 7.4. Биосинтез и обмен аминокислот в растительных организмах. 7.5. Особенности метаболизма белков и аминокислот в животных организмах. 7.6. Биосинтез белка. Нуклеиновые кислоты.. 7.6.1. Механизм биосинтеза белка. 7.7. Вопросы для самоконтроля. 8. химия биологических жидкостей И тканей животных организмов.. 8.1. Биохимия крови. 8.2. Биохимия мяса. 8.3. Биохимия молока и продуктов его микробиальной переработки. 8.4. Вопросы для самоконтроля. 9. БИОХИМИЯ ВИНА.. 9.1. Химический состав винограда и сусла. 9.2. Биохимические процессы на стадии приготовления сусла. 9.5. Вопросы для самоконтроля. литература..
Варианты заданий для самостоятельной работы
2. ВИТАМИНЫ Многие витамины входят в состав клеток животных, растений и микроорганизмов, выполняя одни и те же биохимические функции. Известно более 30 витаминов, их предшественников (провитаминов) и витаминоподобных веществ. Источником витаминов для человека и животных являются растительная пища, а также продукты животного происхождения (молоко, масло, рыбий жир, печень, мясо и др.). При отсутствии витаминов в пище в организме развивается авитаминоз, при недостатке – гиповитаминоз, а при избытке витаминов – гипервитаминоз. Витаминная недостаточность может также развиваться при нарушении обмена веществ вследствие некоторых заболеваний (эндогенная недостаточность). Исторически сложилась тривиальная номенклатура, согласно которой витамины обозначали латинскими буквами (А, В, С, D). В настоящее время используют и систематическую номенклатуру. По физико-химическим свойствам витамины подразделяют на две группы: водорастворимые и жирорастворимые. Жирорастворимые витамины растворяются в неполярных растворителях (нетролинейном эфире, гексане, хлорированных углеводородах, диэтиловом эфире и др.). По биологическому действию на организм витамины делятся на несколько групп (табл. 2.1).
Таблица 2.1 Разделение витаминов на группы
Витамин С представляет собой
Аскорбиновая кислота – сильный восстановитель и легко окисляется даже слабыми окислителями.
Биологическая роль витамина С связывается с участием ее в окислительно-восстановительных процессах. Как восстановитель витамин С – сильный антиоксидант, препятствующий развитию процессов, связанных с окислительной деструкцией биомолекул. Аскорбиновая кислота широко распространена в природе, прежде всего, в фруктах, ягодах и организмах многих животных. В 100 г плодов шиповника содержится до 1000 мг, чёрной смородины – до 560 мг, петрушки – 150 мг, молока – 50 мг, лимона – 40 мг витамина. У людей, обезьян и некоторых других животных витамин С не синтезируется и должен поступать в организм извне.
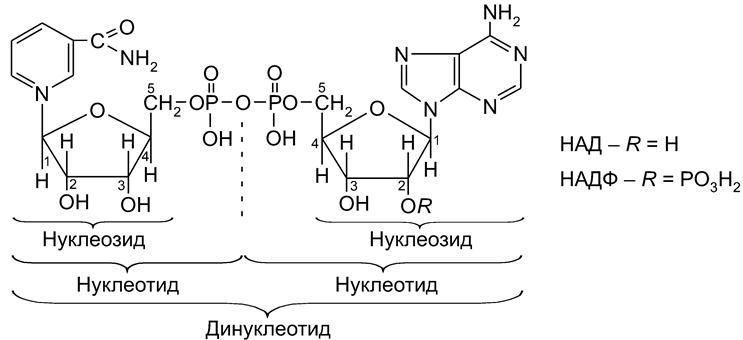
Недостаток в организме витамина РР вызывает заболевание пеллагру (от итальянского pelle agra – шершавая кожа), связанное с болезненными изменениями кожи (дерматитами). Исключительно важная биологическая роль витамина РР определяется тем, что никотинамид является структурным элементом кофермента никотинамид адениндинуклеотида (НАД) и его фосфорилированного по группе ОН производного (НАДФ).
Коферменты НАД и НАДФ являются структурными фрагментами обширного ряда окислительно-восстановительных ферментов дегидрогеназ, относящихся к классу оксидоредуктаз. В частности, эти коферменты “обслуживают” дегидрирование спиртов (в т.ч. полуацеталей) и обратную реакцию – гидрирование карбонильной группы. Естественно, кофементы функционируют, будучи связанными с ферментами – дегидрогеназами. Пиридинзависимые дегидрогеназы относятся к разряду анаэробных дегидрогеназ.
Рибофлавин содержится в продуктах растительного и животного происхождения (в печени, мясе, моркови, пшенице, дрожжах и др.). Рибофлавин входит в состав родственных коферментов – флавинмононуклеотида (ФМН) и флавинадениндинуклеотида (ФАД), участвующих в переносе электронов в окислительно-восстановительных биохимических процессах.
Именно фрагмент изоаллоксазина указанных коферментов переносит молекулу Н2 в цитохромную систему.
В регуляции процесса кровотворения участвует витамин В12. Он обладает лечебным действием при злокачественной анемии у людей. есть соединений, молекулы которых можно формально расчленить на звенья углеродного скелета изопрена (
Витамин А2 отличается от аналога А1 тем, что в цикле содержится две сопряженные двойные связи. Наличие в молекулах каротинов полиеновой системы сопряженных Витамин А – светло-желтая маслянистая жидкость состава С20Н29ОН. Все его двойные связи, кроме находящихся в циклах, имеют транс-конфигурацию. Каротин (С40Н56) – желто-красное вещество. Каротиноиды хорошо растворяются в жирах, легко окисляются, многие из них играют роль витаминов или являются предшественниками таковых. Однако в организмах животных они не синтезируются и должны поступать в них с растительной пищей. В организме Витамин А термостабилен (до 120°С) в безкислородной среде. Под влиянием кислорода быстро окисляется, особенно в кислой среде, чему способствует солнечный свет. Биологическая роль витамина А определяется его участием в зрительном процессе, в ходе которого он окисляется в соответствующий альдегид (ретиналь) и связывается с белком опсином. Витамин А депонируется в печени человека, рыб и животных. Среднесуточная потребность взрослого человека 0,75-1,5 мг. При инфекционных заболеваниях и при работах, связанных с напряжением зрения, доза витамина должна быть увеличена до 4-5 мг. При авитаминозе у человека ослабляется зрение вплоть до полной потери его в ночное время (“куриная слепота”). Авитаминоз у животных выражается в воспалении роговицы и ксерофтальмии, связанной с высыханием и закрытием глаз, ослабляется иммунитет против инфекционных заболеваний, задерживается рост. Прием больших доз витамина А (в 20-30 раз превышающих норму) может привести к отравлению и даже гибели человека. Витамин А частично выделяется с мочой в неизменном состоянии. Главным источником витамина А является каротин, содержащийся вместе со своим ациклическим изомером (ликопином) в моркови, томатах, во многих плодах, в кровяной сыворотке, печени рыб и животных. Витамины группы D встречаются только в организме животных. В растениях содержатся стеролы, из которых при облучении ультрафиолетовыми лучами образуются витамины. По этой причине они называются провитаминами D. Так, в дрожжах и плесневых грибах содержится в большом количестве эргостерол, используемый для промышленного получения витамина D2.
Наиболее распространенной формой витамина D является холекальциферол (витамин D3, получаемый при облучении 7-дегидрохолестерола.
Витамины группы D нерастворимы в воде, но хорошо растворимы в жирах и органических растворителях, в частности, в углеводородных (нетролейном эфире, гексане и др.). Витамины этого ряда устойчивы к высокой температуре, но быстро разрушаются под действием окислителей и минеральных кислот. Реально биологической активностью обладает не сам витамин D3, а продукт его гидроксилирования – 1,25-дигидроксихолекальциферол, который в кишечнике и костях регулирует обмен Са2+ и фосфора. Солнечные лучи способствуют такому превращению в организме. При недостатке в организме витамина D развивается заболевание рахит, связанное с уменьшением содержания кальция и фосфора. При этом размягчаются кости, перерождаются мышцы и ткани. Суточная потребность взрослого человека в витамине D 10-20 мкг. Источники витамина D: печень, рыбий жир, сливочное масло и др. Как и все жирорастворимые витамины, витамин D всасывается в кишечнике и перевариваются вместе с жирами, что требует предварительной эмульгации последних желчными кислотами. Гипервитаминоз опасен преждевременным окостенением и торможением роста детей. Понятие витамин Е объединяет 7 близких по структуре соединения, из которых наиболее важными являются Наибольшей биологической активностью обладает
Токоферолы устойчивы к нагреванию (до 175°С), менее стойки в кислой и щелочной средах. Недостаток витамина Е в кормах приводит к нарушению половой функции у самок и самцов, возникает мышечная слабость (дистрофия), нарушается функция нервной системы и, в целом, обмена веществ. У человека наблюдается дегенерация печени и нарушение функции клеточных мембран. Предполагается, что витамин Е препятствует окислению липидов в структуре клетки и пуриновых оснований. Токоферол – природный антиоксидант. Механизм действия антиоксиданта (будь то в живой клетке, в пищевом продукте или в полимерном материале) заключается в том, что, сталкиваясь с активным радикалом ( Среднесуточная потребность человека в витамине Е – 10-20 мг. Источники витамина: растительные масла, семена пшеницы, яйца, мясо и др. Аналогично ведут себя полиалкилфенолы, многоатомные фенолы и их производные, в том числе пропиловый эфир галловой кислоты (пищевой антиоксидант прогалин Р, или РG), кверцетин и его гликозиды.
Один из глюкозидов кверцетина – рутин – относится к группе витаминов Р. Таннины дубильных веществ представляют собой глюкозу, ацилированную галловой и м-дигалловой кислотами, что сообщает им свойства антиоксиданта. Существует две основные формы витамина – витамин К1 (филлохинон) и витамин К2 (монахинон). Они являются производными бензохинона.
В другой модификации – витамине K2 – боковую цепь составляет шесть изопреновых звеньев, в каждом из которых имеется двойная связь. Витамин К1 – маслянистая жидкость (т.пл. 20°С), чувствителен к свету, разрушается при действии ультрафиолетовых лучей. Витамин К2 кристаллическое вещество (т.пл. 50-52°С). Витамин К (витамин коагуляции) участвует в образовании белка плазмы крови – протромбина – и обеспечивает нормальную свертываемость крови. Витамин К в воде не растворяется, что ограничивало использование его в клинике для борьбы с кровотечением. А.В. Паладин синтезировал растворимый в воде заменитель витамина К – викасол (гидросульфитное производное 2-метил-1,4-нафтохинона).
Витамин К содержится в хлоропластах листьев зеленых растений. Его боковая цепь является остатком высокомолекулярного алифатического спирта фитола, входящего в состав хлорофилла. Витамин К синтезируется микроорганизмами в толстом кишечнике. 2.4. Вопросы для самоконтроля Известно более 2000 различных ферментов, из которых порядка 90% не изучены. Выделены в кристаллическом состоянии или в виде Молекулярная масса ферментов варьируется в пределах 12 000-1 000 000. Двухкомпонентные ферменты являются сложными белками, состоящими из белковой компоненты (апофермента) и небелковой части – кофермента, или простетической группы. Весь комплекс называется холоферментом: Холофермент
Классификация и номенклатура ферментов основывается на природе химических превращений. Выделяют шесть главных классов ферментов: 1. Оксидоредуктазы – катализируют окислительно-восстановительные реакции. 2. Трансферазы – контролируют реакции межмолекулярного переноса групп атомов. 3. Гидролазы – обеспечивают реакции гидролитического расщепления химических связей. 4. Лиазы – катализируют реакции негидролитического расщепления, присоединения групп по двойной связи и обратные реакции отщепления таких групп. 5. Изомеразы – контролируют реакции изомеризации. 6. Лигазы (синтетазы) – катализируют реакции соединения двух молекул, сопряженные с расщеплением связей Р–О в молекулах АТФ или нуклеозидтрифосфата. Классы разделяются на подклассы. Так, оксидоредуктазы классифицируются по природе окисляемой группы в субстрате (спиртовой С–ОН, альдегидной СНО, аминогруппы С–NH2 и др.) Трансферазы классифицируются согласно транспортируемых групп атомов (ацильных, азот-, фосфор-, серосодержащих групп и т.д.). Гидролазы подразделяются по типу гидролизуемых связей (эфирных, пептидных, амидных и др.). Лиазы идентифицируются по природе подвергающихся разрыву связей (С–О-лиазы, С–N-лиазы, С–С-лиазы и др.). Изомеразы различают по типу реакции изомеризации (цис-/транс-изомеризация, рацемизация и др.), а лигазы – по природе вновь образующихся связей. В свою очередь, каждый подкласс делится на подподклассы, названия которых детализируют природу ферментативной реакции. Краткие названия некоторых распространенных групп ферментов и их функции (отмечены в скобках) формулируются следующим образом. 1. Дегидрогеназы (дегидрирование субстратов). 2. Оксидазы (окисление субстратов кислородом). 3. Гидроксилазы (введение группы ОН с помощью О2). 4. Оксигеназы (включение молекулы кислорода при расщеплении С–С-связи). 5. Киназы (перенос фосфатной группы на субстрат от АТФ). 6. Трансферазы (перенос определенных групп). Так, трансацетилаза переносит ацетильный радикал, транскарбоксилаза переносит карбоксильную группу и т.д. Покажем названия ферментов на примерах следующих реакций. 1. Шифр фермента – КФ 1.1.1.1. Систематическое название – алкоголь: НАД-оксидоредуктаза. Тривиальное название – алкогольдегидрогеназа. Ферментативная система работает по принципу “замок-ключ” (к замку подходит только ключ со строго определенной конфигурацией). Геометрические параметры белкового окружения реакционного центра фермента таковы, что в эту нишу укладывается молекула субстрата со строго определенным пространственным строением (матричный эффект). Известна относительная групповая специфичность. Так, липазы и эстеразы расщепляются не только триацилглицеролы (жиры), но и диацилглицеролы, моноацилглицеролы, другие сложные эфиры. А пептидазы расщепляют пептидные связи в различных полипептидах и белках. Особое значение имеет стереохимическая специфичность ферментов. Например, фумарат-гидратаза (L-малат-гидролиаза) катализирует превращение только фумарата, имеющего, как известно, транс-конфигурацию. Хотя существуют и ферменты, действующие как на цис-, так и транс-изомеры. 3.4. Коферменты Можно выделить три группы кофаторов: 1) Специфические коферменты, или коэнзимы – низкомолекулярные органические соединения, обратимо связывающиеся с белками ферментов (апоферментами). 2) Простетические группы – коферменты, прочно связывающиеся с апоферментом, как правило, за счет ковалентных связей. 3) Активаторы – неспецифические вещества, способствующие повышению каталитической активности ферментов. Особое место среди коферментов первой группы в классе оксидоредуктаз занимают коферменты, в состав которых входит ион металла и порфириновое ядро. Магниевый комплекс замещенного порфирина – хлорофилл – принимает участие в фотосинтезе углеводов. А соответствующий комплекс с Fe (ІІ) – гем – служит коферментом для белков (гемоглобина, миоглобина), осуществляющих перенос кислорода в процессе дыхания. Вторую группу коферментов составляют переносчики групп атомов, связанные с ферментами класса трансфераз: нуклеозидфосфаты (аденозинтрифосфат, или АТФ, уридиновые нуклеотиды), кофермент А (КоА). Аденозинтрифосфат (АТФ) является макроэргическим (то есть богатым энергией) аккумулятором биологической энергии:
Переносчиками групп в ферментативных реакциях субстратов, содержащих альдегидную и ацильную группы, являются ЦДФ-холин и ЦДФ-этаноламин. В обмене углеводов важную роль играют уридиннуклеотидные коферменты. В качестве переносчика ацильных фрагментов в реакциях ацилирования и поставщика ацетильного радикала (
Ацильные радикалы локализуются на группе SH кофермента (
4.1. Общая стратегия энергообеспечения организмов 4.2. Переносчики химической энергии и некоторые пути их трансформации 4.3. Анаэробный и аэробный механизм окисления 4.4. Цитохромы и их предшественники. Другие конечные оксидазы 4.5. “Прямые” оксидазы 1. Аденозиндифосфат (АДФ) и аденозинтрифосфат (АТФ)
2. Карбоксилфосфатная связь, например, в 1,3-дифосфоглицериновой кислоте:
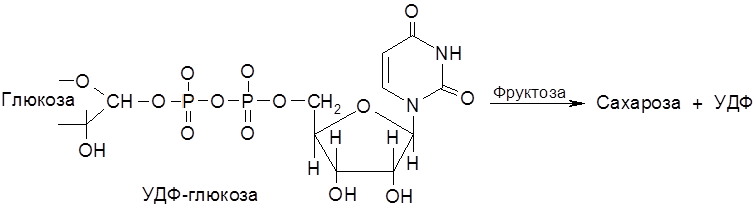
В АТФ под влиянием специфических ферментов могут расщепляться различные связи Р–О. В обмене веществ участвует ряд нуклеозидполифосфатов – моно-, ди- и трифосфаты уридина, гуанидина, цитидина и инозина. Например, синтез сахарозы в растениях осуществляется с участием уридиндифосфатглюкозы (УДФ-глюкозы):
АТФ является своеобразной “разменной монетой” – единицей измерения накопленной и расходованной в процессе биосинтеза энергии. При гидролизе молекулы АТФ до АДФ выделяется 30,6 кДж энергии. Различают субстратное и окислительное фосфорилирование. Субстратное фосфорилирование осуществляется в цитоплазме клетки. Примером субстратного фосфорилирования может служить фосфорилирование АДФ с помощью сукцинил-КоА в растительной клетке:
Окислительное фосфорилирование в дыхательной (электронопроводящей) цепи заключается в высвобождении энергии молекул водорода, локализованных на коферментах (НАД·Н 2, ФАД·Н2) дегидрогеназ. Одна молекула НАД·Н2 продуцирует три молекулы АТФ. Суммарное уравнение окисления НАД·Н2 в дыхательной цепи, сопровождаемого фосфорилированием, можно представить уравнением: НАД·Н2 + 3АДФ + 3Фн + 1/2О2
В живой клетке реализуется два типа ферментативного окисления органических веществ – анаэробный, протекающий без участия кислорода, и аэробный, связанный с окислительным действием на заключительных стадиях кислорода. В основе первого лежат процессы дегидрирования (отщепления молекул Н2) (В.И. Палладин, Г. Виланд, О. Варбург) под влиянием ферментов дегидрогеназ и декарбоксилирования, осуществляемого ферментами декарбоксилазами. Высвобождаемая в этих процессах биоэнергия аккумулируется за счет фосфорилирования АДФ в АТФ. Суть этого процесса заключается в том, что вначале молекула воды внедряется в субстрат, затем ферменты дегидрогеназы отщепляют молекулу водорода, оставляя субстрат в окисленной форме.
Процессы окисления, связанные с дегидрированием, реализуются в условиях анаэробного окисления глюкозы (гликолиз, брожение), при окислительной деструкции пировиноградной кислоты и ацетил-S-КоА ( В процессе аэробного окисления в качестве акцептора (окислителя) молекул водорода на заключительных стадиях функционирует кислород. Существует три способа окислительного действия кислорода: 1. Окисление водорода до воды в дыхательной цепи. 2. Окисление водорода НАД·Н2 и НАДФ·Н2 с участием промежуточных агентов – аэробных дегидрогеназ флавинового типа. Флавопротеин – желтый дыхательный фермент, отщепляющий Н2 от НАДФ·Н2 при окислении гексозомонофосфата в фосфоглюконовую кислоту передает водород коферменту ФАД·Н2 и далее – кислороду воздуха. 3. Третий путь окисления связан с непосредственным воздействием окислителя на субстрат с внедрением в его структуру одного или двух атомов кислорода (А.Н. Бах). Окисление такого типа осуществляется так называемыми “прямыми” оксидазами и не сопровождается образованием АТФ. 4.4. Цитохромы и их предшественники.
Ферменты цитохромы, обозначаемые латинскими буквами (а, а3, в, в3, в6, в7, с, с1, d, f, o и др.), представляют собой протеиды (белки), содержащие в качестве простетических групп гемы. Последние являются порфириновыми комплексами железа. Физиологическая функция цитохромных систем заключается в окислении кислородом молекул водорода, в частности, в цикле трикарбоновых кислот. Цитохромы существуют в окисленной и восстановленной формах, легко превращающихся друг в друга в результате переноса электрона:
В качестве промежуточных звеньев в переносе электронов молекул водорода от восстановленных анаэробных дегидрогеназ к кислороду выступают наряду с цитохромами флавопротеиды (флавинзависимые дегидрогеназы), негемовые железопротеиды (ферредоксины) и жирорастворимые хиноны (убихиноны, или коферменты Q). Процессы аэробного окисления у клеток прокариот локализованы в клеточной мембране, а у эукариот – в митохондриях. Ферментативное окисление молекулярным кислородом катализируется оксигеназами. Некоторые из них выступают в качестве аэробных дегидрогеназ – отщепляя от субстрата водород, они превращают его в воду или пероксид водорода. Другие обеспечивают внедрение в молекулу субстрата двух атомов кислорода, сопряженное в ряде случаев с расщеплением ароматических или алициклических колец. Оксигеназы третьего типа, называемые гидроксилазами, внедряют в субстрат один из двух атомов молекулы кислорода. Типичными представителями оксигеназ являются полифенолоксидазы (фенолоксигеназы).
В растениях широко распространен фермент липооксидаза (липооксигеназа), обусловливающий окисление кислородом воздуха некоторых ненасыщенных длинноцепных жирных кислот и образуемых ими сложных эфиров, в частности, жиров. Липоксигеназа разрушает каротин в процессе сушки и хранения растительных продуктов, а также обусловливает прогоркание масел, круп и муки. К группе “прямых” оксигеназ относится уратоксидаза (уриказа). В растениях содержится оксидаза гликолевой кислоты – гликолатоксидаза:
В плесневых грибах содержится фермент глюкозооксидаза, способная окислять глюкозу в глюконовую кислоту. Ферментами прямого биологического окисления являются каталаза и пероксидаза. Биологическая функция каталазы заключается в разложении пероксида водорода:
Пероксидаза катализирует окисление субстратов (
Пероксидаза содержится в слюне, соке поджелудочной железы, печени, лейкоцитах, в корне хрена, в млечном соке растений. Выделена и охарактеризована пероксидаза хрена (гемопротеид, М=40,000).
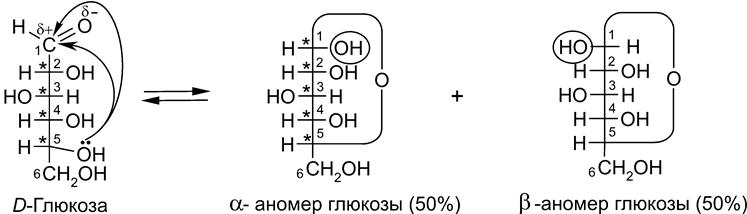
5.1. Наиболее важные представители – строение и свойства. 5.2. Обмен углеводов – анаболизм и катаболизм. Брожение и аэробное окисление. 5.2.1. Гликолиз. 5.2.2. Пировиноградная кислота. 5.2.3. Цикл трикарбоновых кислот (ЦТК, цикл Кребса). 5.2.4. Глиоксилатный цикл. 5.2.5. Пентозофосфатное (апотомическое) окисление моносахаридов. 5.2.6. Особенности обмена углеводов в организмах животных и человека. 5.2.7. Аэробный распад (брожение) углеводов (сахаров). 5.2.8. Фотосинтез. 5.2.9. Взаимопревращения углеводов в растительных организмах. 5.2.10. Хемосинтез. 5.1. Наиболее важные представители – Свойства моноз обусловлены наличием гидроксильных и карбонильной (альдегидной или кетонной) групп. В частности, гидроксильную группу, как носитель нуклеофильных свойств, можно алкилировать, ацилировать, замещать на галоген и аминогруппу. Кето-енольная таутомерия обусловливает взаимное превращение в растворе представленных выше глицеринового альдегида и диоксикетона, а также глюкозы, фруктозы и маннозы. Моноза может обратимо присоединить к оксогруппе свою собственную спиртовую группу с образованием циклического полуацеталя ( кольчато-цепная таутомерия).
Гидроксил , возникающий вследствие циклизации молекулы моносахарида на основе карбонильного кислорода (обведен кружком), называется полуацетальным, или гликозидным (в случае глюкозы – глюкозидным). Далее полуацетальный гидроксил может легко замещаться на алкоксильный радикал с образованием ацеталя. Продукты замещения атома водорода гликозидного гидроксила на алкильный радикал (в т.ч. и несущий различные функциональные группы) получили название гликозидов. Сам радикал при этом называется агликоном. Атака одной из гидроксильных групп монозы на собственную карбонильную группу равновероятна с обеих сторон, поэтому в этой реакции образуются два изомера (диастереомера) (50х50%), отличающиеся лишь конфигурацией атома углерода исходной карбонильной группы. Эти диастереомеры называются аномерами и обозначаются как α- и β-формы (α- и β-аномеры). Положение заместителей в монозах указывается с помощью формул (проекций) Фишера (1) или перспективных формул Хеуорса (2):
D- Глюкоза (виноградный сахар) содержится в винограде, фруктовых плодах и корневищах растений, входит в состав меда, содержится в организмах животных (в крови, лимфе, спинномозговой жидкости и т.д.). Фруктоза – промежуточный продукт метаболизма глюкозы (гликолиза) и гликогена – животного крахмала (гликогенолиза). D -(+)-Галактоза (цереброза) входит в состав дисахарида лактозы (молочного сахара), трисахарида раффинозы, полисахарида агар-агар, гуммиарабика. D -(+)-Манноза входит в состав сложных углеводов, обнаруживается в сыворотке крови, белках слюны, слизи кишечника. Манноза сбраживается дрожжами. D- Рибоза и D-дезоксирибоза входят в состав соответственно рибонуклеиновых и дезоксирибонуклеиновых кислот (РНК и ДНК):
К олигосахаридам относятся ди-, три-, тетра- и другие низкомолекулярные полисахариды, содержащие до 10 звеньев моносахаридов. По существу олигосахариды это простые полиэфиры, в которых связи C–O–C (гликозидные связи) образуются за счет отщепления молекулы воды от двух гидроксильных групп – полуацетальных (гликозидных) или спиртовых (в гексозах – чаще всего в положениях 4 или 6) по схеме:
Дисахариды (биозы) можно разделить на две основные группы: 1. Гликозидогликозиды, не дающие реакций на оксогруппу 2. Гликозидомонозы, дающие указанные реакции на карбонильную группу в соотношении 1 моль реагента на 1 моль дисахарида, то есть с участием одной половины молекулы (такие сахара относятся к типу редуцирующих). Сахароза – это один из самых распространенных в природе дисахаридов. Он содержится в листьях, стеблях и корнях растений, в семенах, в ягодах и фруктах. В частности, в корнеплодах сахарной свеклы содержится до 29% сахара, а в стеблях сахарного тростника – около 20%.
Мальтоза в свободном виде не встречается, образуется при неполном гидролизе крахмала под влиянием фермента диастазы, является промежуточным продуктом в винокуренной и пивоваренной промышленности. Полисахариды представляют собой природные высокомолекулярные вещества (молекулярная масса – от 20.000 до 10.000.000 и выше), построенные по типу биоз. При полном их гидролизе в кислой среде образуются монозы. Полисахариды, образующие при этом монозу одного типа (крахмал, клетчатка, инулин и др.), называются гомополисахаридами, а состоящие из двух или более типов моноз – гетерополисахаридами. Гомополисахариды: клетчатка, крахмал, гликоген – состоят из остатков глюкозы; инулин построен из блоков фруктозы. К гетерополисахаридам отнясятся гемицеллюлозы (полуцеллюлозы) – полимеры растительного происхождения, имеющие разветвленную структуру. Одним из важнейших гомополисахаридов является крахмал. Крахмал – запасной углевод растений – депонируется в клубнях картофеля, зернах злаков и многих семенах. Он составляет весомую составную часть пищи человека. Содержание его в рисе – 78%, в муке – 74%, в белом хлебе – 51%. Полисахариды крахмала состоят из двух фракций – амилозы (5-25%) и амилопектина (75-95%). Глюкозные остатки в неразветвленных частях полисахаридов крахмала имеют конформацию ванны и соединяются α-1,4-глюкозид-глюкозными связями:
или Ваннообразная конформация α-D-глюкопиранозных звеньев способствует спирализации полиглюкозидной цепи, фиксируемой водородными связями. По этой причине крахмал имеет форму микроскопических зерен. Гликоген – полисахарид, содержащийся в тканях тел человека и животных, в грибах, дрожжах, в зерне сахарной кукурузы. Целлюлоза (клетчатка) – нативные ВМС, содержащие до 10.000 звеньев глюкозы с молекулярной массой 500.000-20 млн. – главный конструкционный материал растительных клеток. Достаточно чистая (до 90%) природная клетчатка составляет основу волокна хлопка, ваты, фильтровальной бумаги. Высокая прочность целлюлозы объясняется ее структурой. Полимерные цепи целлюлозы составляют кольца β-глюкозы в форме кресла, что обусловливает их линейное расположение, исключая возможность спирализации: Инулин – полисахарид, состоящий на 94-87% из остатков молекул β-фруктозы, соединенных за счет гликозидных гидроксилов, является резервным энергетическим материалом многих растений (цикория, георгин, кок-сагыза и др.). М = 5000-6000. В растениях, плесневых грибах и дрожжах содержится особый фермент – инулаза, который расщепляет инулин с образованием фруктозы. Инулин сладок на вкус, используется в качестве заменителя сахарозы и крахмала при сахарном диабете.
В фруктах, ягодах, клубнях и стеблях растении обнаруживаются пектины – полимерные вещества, макромолекулы которых состоят из соединенных 1,4-гликозидными связями: остатков галактуроновой кислоты и ее метиловых эфиров (степень метоксилирования – 30-80%) (см. п. 12.1.1).
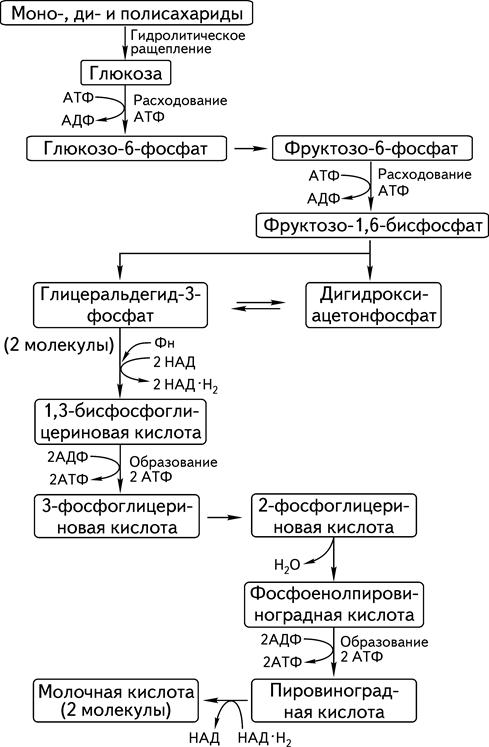
1. Изомеризация глюкозы во фруктозу и расщепление последней на две таутомерные триозы: глицеральдегид и диоксиацетон. 2. Окисление глицеральдегида в пировиноградную кислоту (пируват) с последующим восстановлением последней в ряде случаев в молочную кислоту (лактат). Первая стадия начинается с ферментативного фосфорилирования гексоз и расщепления фруктозо-1,6-дифосфата на две триозы. Дигидроксиацетон термодинамически более стабилен, чем глицеральдегид, что объясняет их соотношение в равновесной смеси. Однако именно глицеральдегид продолжает цепь метаболических превращений во второй стадии гликолиза, связанной с окислением его до пировиноградной, а в ряде случаев – до молочной кислоты. Гликолиз в общих чертах представлен на схеме 5.1.
Схема 5.1. Общая схема гликолиза 5.2.2. Пировиноградная кислотаПировиноградная кислота занимает центральное место во всем разнообразии анаэробных и аэробных процессов катаболизма углеводов (схема 5.2).
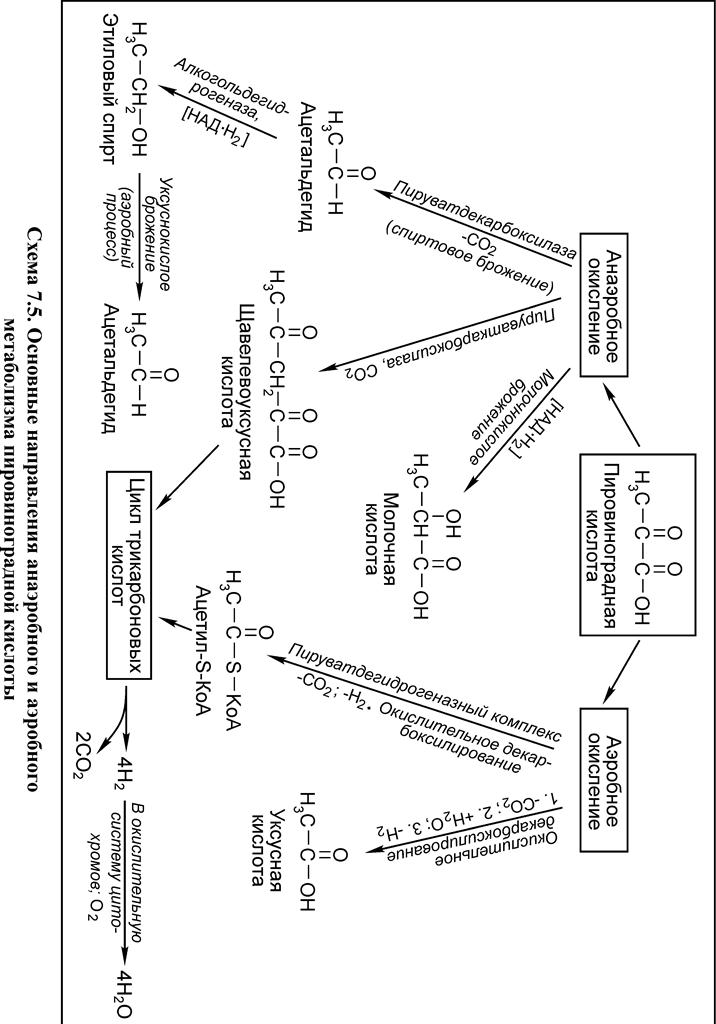
Во второй стадии аэробного окисления пировиноградная кислота подвергается окислительному декарбоксилированию с образованием ацилированного по группе SH кофермента А – ацетил-S-KoA (схема 5.3).
Схема 5.3. Окислительное декарбоксилирование пирувата Высокой энергоемкостью характеризуется третья стадия аэробного окисления глюкозы – диссимиляция двух молей ацетил-S-КоА в цикле трикарбоновых кислот (ЦТК, или цикле Кребса, или цикле лимонной кислоты) (схема 5.4). В общем виде материальный баланс превращений молекулы
Общий энергетический выход аэробного окисления глюкозы до СО2 и Н2О – 38 высокоэнергетических связей АТФ.
Схема 5.4. Цикл трикарбоновых кислот (ЦТК). Ферменты (постадийно): 1 – цитратсинтетаза; 2 – аконитаза; 3 – гидратаза; 4 – НАД-зависимая изоцитратдегидрогеназа; 5 – декарбоксилаза; 6 – оксоглутаратдегидрогеназный комплекс; 7 – сукцинатдегидрогеназа, ФАД; 8 – фумараза; 9 – малатдегидрогеназа. 5.2.4. Глиоксилатный циклСуществуют и другие энергопродуцирующие пути дыхания. Так, некоторые бактерии (Escherichia coli, Pseudomonas), плесневые грибы и растения в качестве источника углерода используют метоболизм уксусной кислоты (ацетата) в так называемом цикле глиоксиловой кислоты.
У жвачных животных гидролитическое расщепление углеводов, в том числе целлюлозы (клетчатки), происходит в значительной мере под влиянием заселяющих рубец (отдел желудка) бактериальных ферментов: целлюлазы, целлобиазы, мальтазы и сахаразы. Образующаяся из целлюлозы целлобиоза, а затем У большинства животных и человека переваривание углеводов происходит главным образом в кишечнике. В клетках печени и мышц часть глюкозы депонируется, превращаясь в гликоген (животный крахмал). 5.2.7. Анаэробный распад (брожение) углеводов (сахаров) Гликолиз может иметь продолжение и в анаэробном процессе – брожении сахаристых веществ. Этот процесс осуществляется благодаря жизнедеятельности ряда микроорганизмов – бактерий и плесневых грибов. Аналогичные процессы, именуемые анаэробным (интрамолекулярным) дыханием, осуществляются в клетках растительных организмов (прежде всего, в плодах и семенах), где они сочетаются с процессами аэробного дыхания. Вклад каждого из этих двух взаимодополняющих типов дыхания зависит от природы организма и внешних условий (газового режима, температуры, влажности и др.). Брожению подвергается ряд моно-, ди- и полисахаридов, но центральное место в этом ряду занимает глюкоза, а из всех видов брожения наиболее важным представляется спиртовое брожение:
Образующаяся на стадии гликолиза пировиноградная кислота подвергается декарбоксилированию с последующим восстановлением ацетальдегида в спирт.
Наряду с главными продуктами спиртового брожения (C2H5OH, CO2) в процессе образуются в незначительных количествах янтарная кислота, сивушные масла (смесь амилового, изоамилового, бутилового и др. спиртов), уксусный альдегид, глицерин и некоторые другие соединения. Наиболее легко сбраживаются глюкоза и фруктоза, медленнее – манноза, еще медленнее – галактоза. Из дисахаридов подходящими субстратами брожения являются сахароза и мальтоза. При молочнокислом брожении молекула гексозы распадается на две молекулы молочной кислоты:
Такое брожение занимает важное место в производстве молочнокислых продуктов (простокваши, кефира, кумыса, ацедофилина), при изготовлении кваса, хлебных заквасок, при квашении капусты, огурцов и при силосовании кормов. Молочная и уксусная кислоты образуются при сбраживании ржаного теста. Совместное существование (симбиоз) дрожжей и молочнокислых бактерий, оказывающих друг на друга благотворное влияние, наблюдается во многих пищевых продуктах и полуфабрикатах. Молочная кислота широко применяется в пищевой, текстильной и кожевенной промышленности. В этой связи представляют интерес высокопроизводительные (по выходу молочной кислоты) термофильные молочнокислые бактерии, подобные широко используемым Termobacterium cereale. Третьим видом брожений является маслянокислое брожение:
Дыхание растительных организмов сопровождается рядом типичных явлений: уменьшение биомассы, изменение состава окружающей среды, усиленное выделение тепла и влаги. 5.2.8. Фотосинтез Фотосинтез – это процесс образования в организмах органических веществ, на основе оксида углерода (IV) и водорода, продуцируемого фотолитическим разложением воды. Суммарно процесс фотосинтеза можно выразить уравнением:
Чаще всего n=6, что соответствует образованию гексоз:
Фотосинтез – это основной источник органических соединений и молекулярного кислорода на Земле. Хлорофиллы являются комплексными соединениями – порфиринатами магния (II), обнаруживающими сходство с порфиринатами железа (II) (гемами).
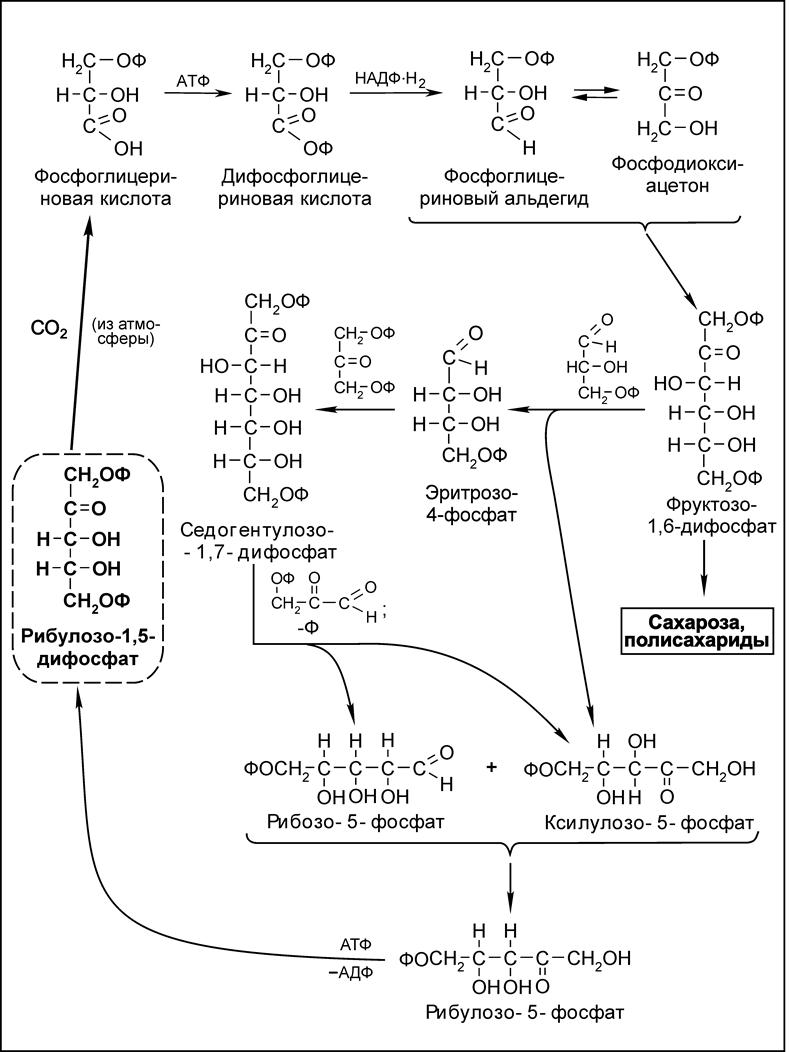
Далее процесс фотосинтеза находит продолжение в темновой фазе. Здесь образовавшиеся 2 молекулы НАДФ·Н2 и 3 молекулы АТФ используются для восстановления одной молекулы оксида углерода (IV) до углевода. Характер биохимических превращений при фотосинтезе отражает цикл Кальвина (схема 5.6).
Схема 5.6. Биохимические превращения оксида углерода (IV) и рибулозо-1,5-дифосфата в условиях фотосинтеза (цикл Кальвина).
Суммарное уравнение цикла Кальвина:
5.2.9. Хемосинтез Существуют микроорганизмы, не содержащие хлорофилла, которые усваивают СО2 и синтезируют органические вещества, используя энергию, образующуюся при окислении различных неорганических соединений: H2S, S8, H2, NH3, HNO2, производных Fe и Mn. Такой процесс получил название хемосинтеза. Бесцветные серобактерии некоторых водоемов получают энергию для хемосинтеза по реакции:
В почвах и водоемах широко распространены нитрофицирующие бактерии, которые извлекают энергию, необходимую для синтеза органических соединений, путем окисления аммиака и азотистой кислоты. Одним из источников аммиака является гниение белков. 5.3. Вопросы для самоконтроля17. Дайте определение и проведите классификацию углеводов. 18. Покажите типы таутомерии, свойственные моносахаридам. Как отличаются по структуре крахмал и клетчатка? 19. Назовите основные стадии анаэробного дыхания (брожения) и аэробного дыхания. 20. Покажите химизм гликолиза. 21. В чем заключается центральная роль пировиноградной кислоты во всей совокупности анаэробных и аэробных процессов? 22. Из каких углеводов можно получить этанол методом брожения и какие операции следует провести при использовании полисахаридов? 23. Представьте химизм и биологическую роль пентозофосфатного окисления. 24. Какая взаимосвязь существует между аэробным и анаэробным процессами в растительных организмах? 25. Какие особенности обмена углеводов в организмах животных и человека Вам известны? 26. Охарактеризуйте фотосинтез как источник всех соединений углерода на Земле. 6. Липиды – химическое строение, свойства и биологическая роль. кАТАБОЛИЗМ И АНАБОЛИЗМ
Жиры, содержащие много двойных (олефиновых) связей при комнатной температуре имеют жидкую консистенцию и называются маслами (табл. 6.1). Таблица 6.1 Содержание жиров в семенах и плодах
В жирах животного происхождения повышено содержание насыщенных кислот (табл. 6.2), что обусловливает их твердую консистенцию при комнатной температуре. Таблица 6.2 Содержание основных жирных кислот
Таблица 6.3 Содержание жирных кислот в некоторых
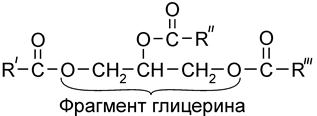
6.2. Фосфолипиды (фосфоглицериды) Фосфолипиды – это глицеролы, содержащие наряду с гидрофобными ацильными группами жирных кислот гидрофильные компоненты фосфорной кислоты и ее производных, включающих остатки спиртов, содержащих аминогруппу (холин НОСН2СН2
6.3. Сфинголипиды Сфинголипиды – это сложные эфиры сфингозиновых оснований (аминоспиртов), наиболее типичным представителем которых является сфингозин
Структурной основой сфинголипидов являются церамиды –
6.4. Стериды и стерины К липоидным соединениям относят группу стеридов – сложных эфиров жирных кислот и стеринов. Последние представляют собой циклические спирты, производные циклопентанпергидрофенантрены.
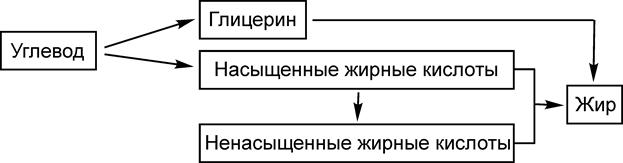
Жиры при длительном хранении прогоркают, приобретают неприятный вкус и запах. Этот процесс может быть обусловлен как чисто химическими реакциями, связанными с действием света, кислорода воздуха и воды, так и действием окислительных ферментов, например, липоксидазы. 6.6. Обмен липидов Схема синтеза жира из углеводов представляется следующим образом:
Главным источником компонентов жира являются глюкоза и фруктоза. Метаболическая цепь превращений, связанных с биосинтезом жирных кислот, начинается со взаимодействия ацетилированного по группе SH кофермента А (
На следующей стадии образовавшийся бутирил-S-АПБ ацилирует новую молекулу малонил-S-АПБ. Далее снова реализуется декарбоксилирование Суммарное уравнение синтеза, например, пальмитиновой кислоты отражает участие семи молекул малонил-КоА.
Диссимиляция (катаболизм) жиров в растительных тканях наиболее интенсивно протекает в прорастающих семенах масличных культур. Первоначальная стадия – гидролиз глицеролов – протекает под действием фермента липазы:
Известно три типа окисления жирных кислот:
Образовавшийся ацилкофермент А подвергается окислительной деструкции по той же схеме. 6.7. Вопросы для самоконтроля 27. Дайте определение липидов и проведите их классификацию. 28. Почему жиры при хранении прогоркают? В чем суть автоокисления жиров? 29. В чем суть биологического окисления жирных кислот. Энергетическая функция кислот. Охарактеризуйте 30. Представьте механизм синтеза жирных кислот на основе малонил-КоА. 7. Белки и аминокислоты – строение,
7.1. Общая характеристика. Биологическая роль 7.2. Аминокислоты – строение и свойства 7.3. Структура и классификация белков 7.4. Биосинтез и обмен аминокислот в растительных организмах 7.5. Особенности метаболизма белков и аминокислот в животных организмах 7.6. Биосинтез белка. Нуклеиновые кислоты 7.7. Пути видоизменения ДНК: мутации, лизогения, трансиндукция, трансформация. Рекомбинация ДНК 7.1. Общая характеристика. Биологическая роль Аминокислоты и продукты их конденсации – полипептиды и белки – являются “краеугольным камнем” “здания” живой природы. В организмах животных белки составляют 14-23% свежей ткани и 45-80% сухой массы. Они являются структурными элементами мышц и тканей, внутренних органов и крови, волос и перьев, входят в состав ферментов, гормонов, антител и большого множества физиологически активных веществ. Источником белка для человека и животных являются пищевые продукты растительного и животного происхождения. Некоторые из них представлены в таблице 7.1. Таблица 7.1 Содержание белка в пищевых продуктах (%)
7.2. Аминокислоты – строение и свойства Основу белков составляют 20 аминокислот. Некоторые из них содержат кроме карбокси- и аминогрупп, другие функциональные группы (–OH, –SH, =NH и др.), ароматические ядра, азотсодержащие гетероциклы. Примеры:
Все природные аминокислоты, кроме аминоуксусной, содержат асимметрический атом углерода и относятся к L-ряду. Аминокислоты обладают всеми свойствами амино- и карбоксильных групп, в частности, их можно алкилировать и ацилировать. Важнейшим свойством аминокислот является ацилирование аминогруппы одной молекулы за счет карбоксильной группы второй молекулы.
Именно такого типа реакции лежат в основе поликонденсации ряда аминокислот в определенной последовательности, ведущей к образованию полипептидов и белков.
Белки и пептиды попадают из желудка в тонкий кишечник, где под действием протеиназах подергаются более глубокому гидролизу. Не гидролизированные в тонком кишечнике белки поступают в толстый кишечник, где они подвергаются гниению. В этом процессе ферменты бактерий осуществляют расщепление белков и аминокислот, сопровождающееся образованием газов (метана, СО2, Н 2S) и токсических веществ (фенола, крезола, скатола, индола и др.). Механизм всасывания аминокислот и низкомолекулярных пептидов связан с их транслокализацией через мембраны клеток с помощью специфических переносчиков. Превращения белков внутри клеток и в межклеточном пространстве органов и тканей осуществляются, как уже отмечалось, комплексом пептидгидролаз ( катепсинами), локализированным в цитоплазме либо на мембранах. Белки в организме постоянно обновляются. Так, белки печени обновляются наполовину за 8-12 суток, белки плазмы крови – за Взаимопревращения аминокислот в процессе обмена в животных организмах основаны на реакциях трансаминирования, дезаминирования (в том числе – окислительного дезаминирования), декарбоксилирования и др. Известен ряд форм выводимого из организма азота: аммиак, мочевина, креатинин, индикан, скатол и индол. Фенол, крезол, индол и скатол – ядовитые вещества с неприятным запахом – всасываются в кровь и при избытке оказывают вредное воздействие на организм. В животных организмах от аминокислот берут начало белки, в том числе ферменты, гормоны, антитела, пиримидиновые и пуриновые основания, необходимые для синтеза нуклеотидов и нуклеиновых кислот, пиррольные производные (порфирины), множество биологически активных веществ пептидной природы. 7.6. Биосинтез белка. Нуклеиновые кислоты Современная научная концепция биосинтеза белков тесно связана с такими проблемами, как наследственность, естественный отбор, выведение новых форм растительных и животных организмов на основании современных достижений генной инженерии. Синтез белка осуществляется в субклеточных структурах – рибосомах, которые локализуются в цитоплазме, митохондриях и хлоропластах. Процесс образования белковых молекул осуществляется по принципу матричного синтеза с помощью нуклеиновых кислот. Нуклеиновые кислоты (полинуклеотиды) представляют собой продукты конденсации мононуклеотидов, состоящих из трех компонентов:
Соединение, включающее два первых компонента – гетероцикл и пентозу – называется нуклеозидом. Таким образом, нуклеотид – это фосфорный эфир нуклеозида. К нуклеозидному фосфату могут присоединяться еще один или два остатка фосфорной кислоты, образуя соответственно нуклеозидди- и нуклеозидтрифосфаты. В состав нуклеотидов и нуклеиновых кислот включены три пиримидиновых основания (урацил, тимин, цитозин) и два пуриновых основания (аденин и гуанин).
Из пентоз в состав рибонуклеиновой кислоты (РНК) включены рибоза, а в дезоксирибонуклеиновую кислоту – дезоксирибоза. Звено Соединение мононуклеотидов посредством эфирных связей фосфорной кислоты приводит к образованию нуклеиновых кислот, принцип построения которой отражает схема 7.1.
Схема 7.1. Стуктура одноцепочечной нуклеиновой кислоты 7.6.1. Механизм биосинтеза белка В соответствии с матричной теорией биосинтеза белка можно выделить следующие его стадии: транскрипция, рекогниция и трансляция. Транскрипция (от лат. transсriptio – переписывание) заключается в том, что на матрице каждой их двух цепей раскручивающейся двойной спирали ДНК в ядре осуществляется синтез аналога рибонуклеиновой кислоты, именуемого матричной (ядерной) РНК. Информационные РНК отделяются от ядра и переходят в протоплазму, где они локализуются на рибосомах и осуществляют поликонденсацию аминокислот, то есть трансляцию (от лат. translatio – перенесение) кода ДНК на структуру белковых молекул. Однако трансляции предшествует стадия рекогниции (от лат. recognition – узнавание, опознание), суть которой состоит в подготовке к введению в процесс поликонденсации молекул аминокислот в строго определенной последовательности. Включение каждой из 20 аминокислот в полипептидную цепь на стадии трансляции обеспечивает так называемый кодон – триплет, состоящий из трех нуклеотидов в цепи м-РНК. Стадию трансляции матричной (информационной) РНК в полипептидную последовательность аминокислот можно также разделить на 3 стадии: инициацию; элонгацию (удлинение) и терминацию (завершение роста цепочки полимера).
Глубокие генетические изменения могут быть достигнуты путем слияния клеток различных типов. В конце 70-х годов в биохимии был разработан ряд методов создания рекомбинантных ДНК, положивших начало генетической инженерии, которая позволяет направленно конструировать генетический материал и вводить его в живые клетки, прививая им полезные с точки зрения экспериментатора свойства. 7.7. Вопросы для самоконтроля Кровь – это вязкая жидкая ткань, осуществляющая через кровеносную систему связь всех органов и тканей организма, создавая необходимый режим их существования. Кровь выполняет различные физиологические функции: транспортную (доставляет кислород и питательные вещества к тканям, удаляет из тканей углекислый газ и продукты обмена), регуляторную (поддерживает постоянным рН и осмотическое давление), защитную (ее лейкоциты и антитела нейтрализуют возбудителей болезней). Кровь способна свертываться при повреждении сосудов, защищая организм от кровопотерь. Состав крови свидетельствует о нормальных и патологических процессах, происходящих в организме. Кровь состоит из плазмы (55-60%) и взвешенных в ней форменных элементов: эритроцитов – красных кровяных телец (39-44%), лейкоцитов – белых кровяных телец (1%) и тромбоцитов (0,1%). Плотность крови – 1,05-1,06 г/см3 Плазма, лишенная белка фибриногена (0,4%) называется сывороткой крови. Эритроциты содержат 40% плотного остатка, на 90% состоящего из гемоглобина. В их составе имеются также белки стромы, фосфолипиды, стериды, гликопротеиды и ряд ферментов: каталаза, карбоангидраза, холинэстераза, ферменты гликолиза. Лейкоциты образуют небольшой слой над выпавшими в осадок эритроцитами. В отличие от эритроцитов лейкоциты имеют ядро и все другие органеллы, присущие соматическим клеткам. Наличие ядра и рибосом свидетельствуют о том, что в лейкоцитах постоянно синтезируются белки, в том числе и ферменты. Лейкоциты играют важную роль в защите организма. Тромбоциты , или кровяные пластинки играют важную роль в образовании кровяного сгустка при свертывании крови. Плазма крови содержит в своем составе 2,5-8% белков, в том числе фибриногена (0,4%). В крови различных видов животных 30-55% от общего количества сывороточных белков составляют альбулины (М=65.000–70.000, рI 4,9; период полураспада 3,7 суток). Они хорошо растворимы в воде, участвуют в регуляции водно-солевого обмена между кровью и тканями, образуют комплексы с металлами, гормонами, витаминами, токсинами, лекарственными препаратами и другими веществами, обеспечивая их транспорт в организме. Глобулины обозначаются буквами Глобулины хорошо комплексуются с различными веществами крови. Особое место среди
Кровь человека и животных характеризуется постоянным значением рН. У различных видов млекопитающих оно варьируется в пределах 7,2–7,8, у человека рН составляет около 7,4. Постоянство рН обеспечивается буферными системами, состоящими чаще всего из слабой кислоты и ее соли с сильным основанием, либо из слабого основания и его соли с сильной кислотой. В организме функционируют следующие буферы: бикарбонатный, фосфатный, гемоглобиновый и белковый. Важной характеристикой крови является осмотическое давление. Осмотическое давление крови 0,8 МПа. Изотоническими сыворотке крови являются 0,9% (0,15 моль/л) раствор NaCl и 5,4% (0,3 моль/л) раствор глюкозы. Такие растворы, называемые физиологическими, используются для приготовления инъекций в вену лекарственных препаратов. Наряду с транспортом питательных веществ кровь выполняет в организме человека и животных дыхательную функцию, то есть осуществляет перенос кислорода от легких к тканям и углекислоты (СО2) от тканей к легким. Способностью связывать большое количество кислорода обладает красный пигмент эритроцитов – гемоглобин (Нb). Некоторые вещества, в частности, промышленные яды (например, нитробензол, хиноны и др.) обладают способностью окислять гемоглобин в метгемоглобин, превращая Fe2+ в Fe3+. 8.2. Биохимия мяса Собственно мясо представляет собой скелетную мускулатуру животных (мышечную ткань). Наряду с мышечной тканью выделяют жировую, костную и соединительную ткани. Химический состав мяса зависит от вида животного, его возраста, физиологического состояния и др. параметров (таблица 8.1). В липидах мяса преобладают жиры, в том числе на основе олеиновой кислоты (33-47%). Содержание холестерина достигает 50-70 мг, а стеридов – 3-5 мг на 100 г продукта. В ткани мяса включены углеводы: глюкоза, гликоген (животный крахмал), гиалуроновая кислота, хондроитинсульфаты. Кроме белков, липидов и углеводов в мясе содержится ряд других вещества. Таблица 8.1 Химический состав мяса различных животных
Гниение мяса обусловлено разложением органических веществ под влиянием ферментов бактерий. При разложении белков образуются индол, скатол, путресцин, кадаверин, фенол, маркаптаны, аммиак и другие продукты. 8.3. Биохимия молока и продуктов Молоко представляет собой биологическую жидкость, которая образуется в молочной железе самок млекопитающих и предназначена для вскармливания новорожденных детенышей. Вместе с тем, молоко ряда животных и продукты его микробиальной переработки широко используются для питания людей всех возрастов. В молоке – около 200 индивидуальных веществ. Содержание наиболее важных из них показано в таблице 8.3. Молоко – жидкость белого цвета, обладающая свойствами полидисперсной системы с элементами истинных и коллоидных растворов, суспензий и эмульсий. Основным углеводом молока является лактоза, выполняющая главным образом энергетическую функцию. Молоко содержит практически все витамины, необходимые для нормального развития человека. Они попадают в него вместе с кормом животных и синтезируются микрофлорой рубца. В молоко из крови поступают гормоны – химические стимуляторы, образующиеся в клетках желез внутренней секреции и регулирующие обмен веществ в организме. В производстве кисломолочных продуктов главное место занимают процессы брожения сахаров, коагуляции казеина и гелеобразования. Брожение молочного сахара (лактозы) осуществляется микроорганизмами бактериальных заквасок. Его скорость и направление определяют консистенцию, вкус и запах готовых продуктов. Таблица 8.3 Химический состав молока (%) некоторых млекопитающих
Примечание : 4,19кДж = 1 ккал. Выделяют две группы кисломолочных продуктов. В основе приготовления продуктов первой группы: простокваши, йогурта, ацидофилина, творога, сметаны – лежит главным образом молочнокислое брожение. Ко второй группе относятся продукты, полученные на основе смешанного брожения – молочнокислого и спиртового: кефир, кумыс, ацидофильно-дрожжевое молоко.
8.4. Вопросы для самоконтроля 9.1. Химический состав винограда и сусла 9.2. Биохимические процессы на стадии приготовления сусла 9.3. Дрожжевое брожение виноградного сусла
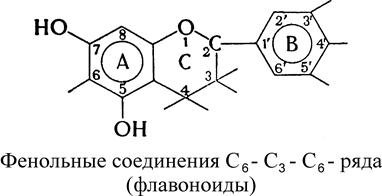
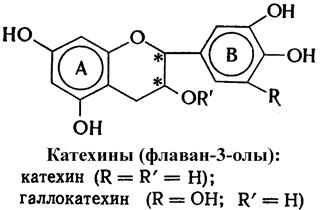
В состав винограда и вина включены химические соединения различных классов: углеводы, органические и ароматические кислоты, фенолы и их гидроароматические производные, азотистые, минеральные и другие вещества. Биохимия вина изучает сущность ферментативных процессов, происходящих при изготовлении вина. Знание механизма ферментативных реакций на стадиях формирования и образования вина позволяет управлять технологическими процессами. 9.1. Химический состав винограда и сусла В ягодах винограда содержатся сахара, главным образом глюкоза, фруктоза и сахароза, органические кислоты, ферменты, витамины, азотистые и фенольные вещества – всего более 400 компонентов. Самыми массоемкими представителями моносахаридов в ягодах винограда являются D-глюкоза и D-фруктоза (10-30% и более). В основном фотосинтез углеводов виноградной лозы осуществляется в зеленых листьях. В начале формирования ягод винограда содержание моносахаридов составляет около 1% и представлено в основном глюкозой. Фруктоза появляется позже, и в период налива ягоды соотношение глюкозы к фруктозе приближается к единице. Большинство штамов дрожжей предпочтительнее сбраживают глюкозу, хотя отдельные виды, например, сотернские дрожжи, лучше сбраживают фруктозу. Из ряда олигосахаридов, включающих до 12 звеньев моносахаридов (полисахаридов I порядка), в винограде больше всего содержится сахарозы. В незначительных количествах присутствуют также мальтоза, целлобиоза и трисахарид рафиноза. Азотистые вещества винограда состоят из органических и минеральных форм азота. К первым относятся белки, полипептиды, аминокислоты, амиды и амины, а ко вторым – нитраты, нитриты и аммиачные соли (всего 2,3-10,0 г/л сока). Основная доля азота приходится на аминокислоты и полипептиды (38-78%). Минеральные формы азота составляют 5-15%. В ягодах винограда содержится значительное количество органических кислот, представленных главным образом винной и яблочной кислотами. В небольших количествах присутствуют также уксусная, лимонная, янтарная, гликолевая, щавелевая и др. кислоты. Благодаря создаваемой ими кислотности в сусле подавляется развитие болезнетворных микроорганизмов и создаются благоприятные условия для деятельности винных дрожжей. В растениях, в том числе в винограде, встречаются разнообразные фенольные соединения моно-, олиго- и полимерного характера. Мономерные фенольные соединения подразделяются на соединения С6–С1-, С6–С3- и С6–С3–С 6-рядов. Например:
Виноград содержит эфирные масла, придающие ему своеобразный аромат. В основном они сосредоточены в кожице и во внешних слоях мякоти винограда. При переработке винограда эфирные масла переходят в сусло, а затем в вино и участвуют в формировании букета напитка. 9.2. Биохимические процессы на стадии На стадии разрушения растительных клеток виноградный сок и мезга насыщаются кислородом. Ягоды винограда содержат ферментативные окислительные системы. Все это обусловливает прохождение окислительных процессов. При выдержке сусла в аэробных условиях в течение 12 ч. содержание фенольных соединений заметно уменьшается, а количество хинонов увеличивается и достигает 10 мг/л. По сравнению с катехинами антоцианы и антоцианиды окисляются в меньшей степени. Виноделие является биотехнологическим производством, связанным с жизнедеятельностью дрожжей, в основе которой лежат биохимические превращения углеводов (сахаров) и продуктов их первичного метаболизма. Основную роль в обмене веществ дрожжевой клетки играют анаэробный и аэробный распад углеводов. При сравнении бродильных свойств рас винных (S. vini) спиртовых (S. cerevisiae) и пивоваренных (S. carlsbergensis) дрожжей установлено, что винные дрожжи наряду с большей спиртоустойчивостью и способностью сбраживать более высокие концентрации сахаров, нуждаются в относительно большем доступе воздуха. Причем, в ряду видов дрожжей, используемых в виноделии, дрожжи S. vini обладают наиболее высокой степенью анаэробиоза. Виноградные ягоды ко времени созревания заселяются не только дрожжами, но и различными видами бактерий. Однако развитие бактерий в сусле и вине сильно подавляются в связи с высокой кислотностью и осмотическим давлением сахаров. Действие осмотического давления обусловлено тем, что высокая концентрация сахаров в сусле способствует обезвоживанию клеток бактерий. Бактерицидное действие вина усиливается с повышением концентрации спирта и рН среды. Бактерии туберкулеза, тифа, холерные вибрионы и кишечная палочка погибают в нем, причем, в белом вине гораздо быстрее (за 15-20 мин.), чем в красном (2-4 час.). В вине могут развиваться только бактерии, выдерживающие рН 2,5-4,5 и концентрацию спирта 12-18%. К ним относятся некоторые виды молочно-кислых бактерий, вызывающих молочнокислое, яблочно-молочное и маннитное брожение, а также отдельные штамы уксуснокислых бактерий. Молочно-кислые бактерии делятся на гомоферментативные (L.plantarum, L.casei и др.) и гетероферментативные (палочки –L.вBuchneri, L. вrevis. L. hilgardii; кокки – Leuconostos mesenteroides, Leuc. Citrovorum, Leuc. vini, Leuc. gracile и др.). Молочнокислому брожению подвергаются все типы вин: сухие столовые, полусладкие, десертные и в особенности крепкие, а также яблочные вина. Типичные гомоферментативные бактерии превращают около 95% глюкозы в молочную кислоту обычным путем гликолиза.
Гетероферментативное молочнокислое брожение выражается общей схемой:
Пировиноградная кислота частично декарбоксилируется с образованием уксусного альдегида. В результате превращений уксусного альдегида и пировиноградной кислоты образуется янтарная и уксусная кислоты. Важным, а по мнению некоторых специалистов единственно полезным процессом, который осуществляется молочнокислыми бактериями в вине, является яблочно-молочное брожение (ЯМБ), вызывающее понижение кислотности в высококислотных винах. Превращение двухосновной яблочной кислоты в одноосновную – молочную кислоту снижает кислотность вина (рН повышается). Уксуснокислые бактерии вызывают соответствующее брожение, суть которого заключается в окислении этанола в уксусную кислоту.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
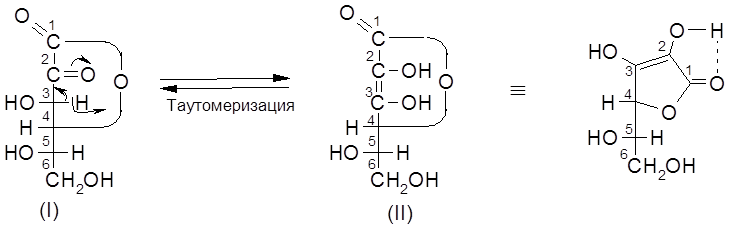
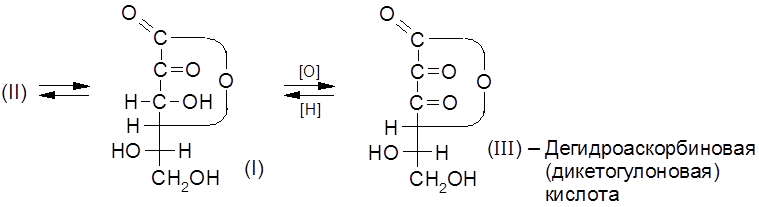
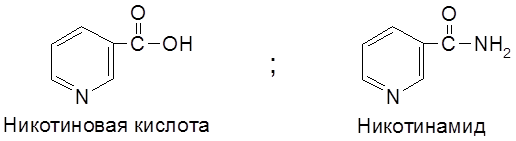
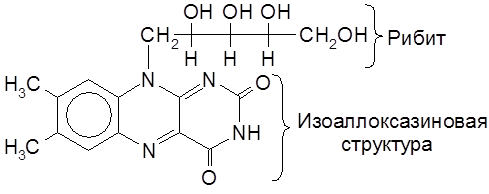
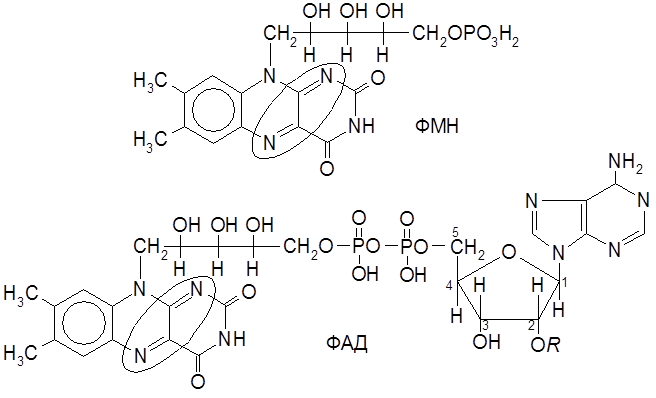
 ). К этой группе относится обширный ряд растительных пигментов – каротинов (каротиноидов) от лат. carota – морковь), из которых наибольшее значение имеют витамин А1 (ретинол), выделенный из печени морских рыб, витамин А2 (дегидроретинол), полученный из печени пресноводных рыб и
). К этой группе относится обширный ряд растительных пигментов – каротинов (каротиноидов) от лат. carota – морковь), из которых наибольшее значение имеют витамин А1 (ретинол), выделенный из печени морских рыб, витамин А2 (дегидроретинол), полученный из печени пресноводных рыб и 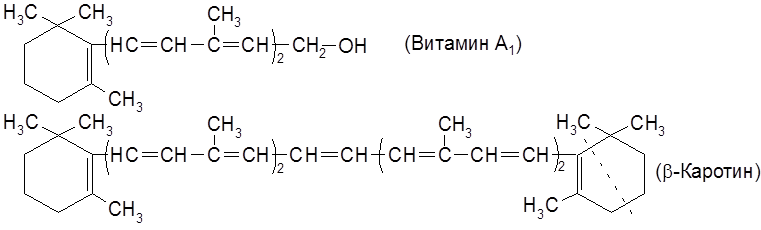
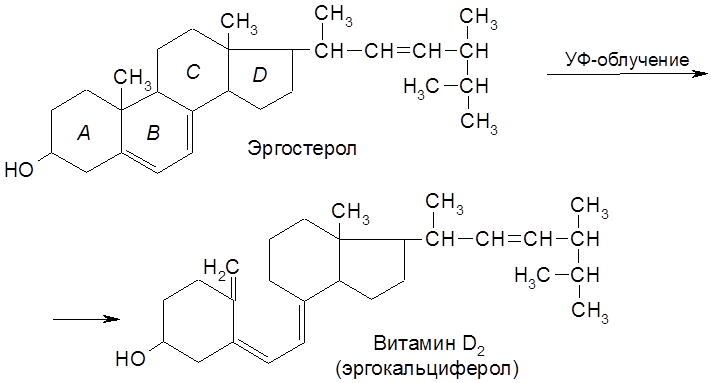
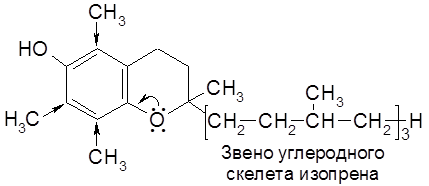
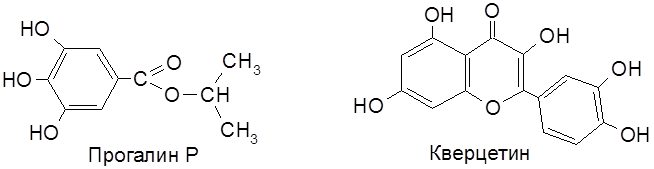
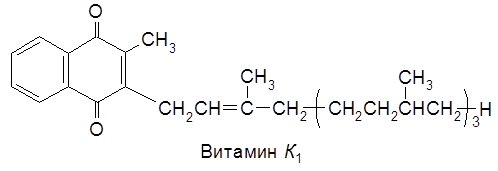
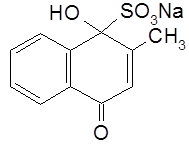
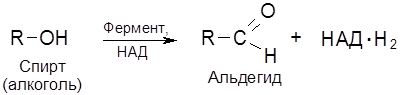
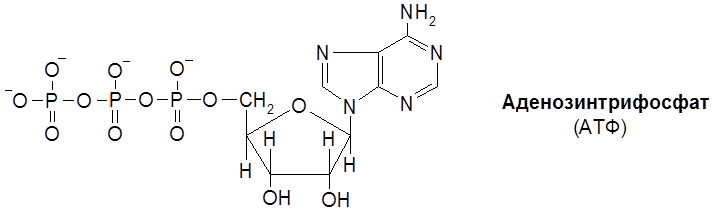
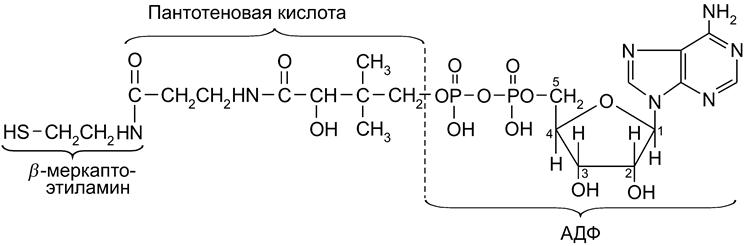
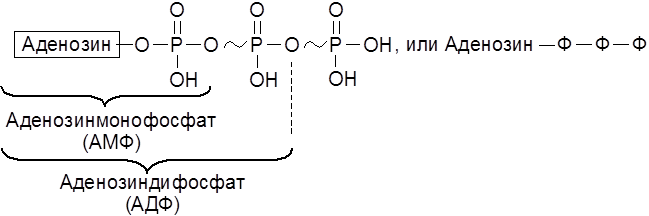
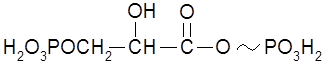
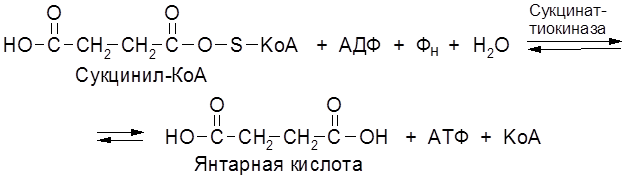
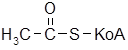 ) в цикле трикарбоновых кислот (ЦТК), в пентозофосфатном окислении глюкозы, в реакциях
) в цикле трикарбоновых кислот (ЦТК), в пентозофосфатном окислении глюкозы, в реакциях 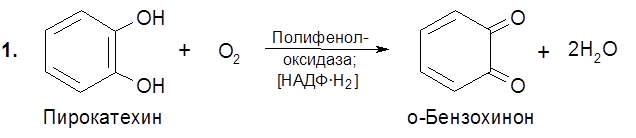
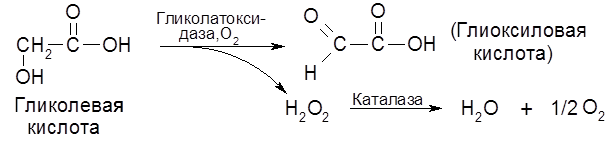
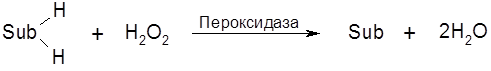
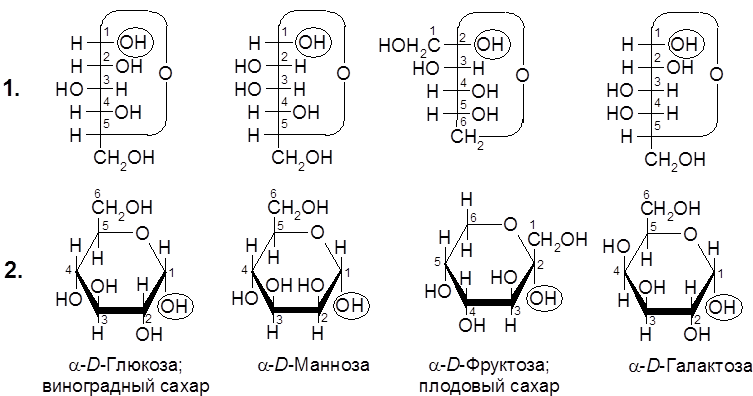

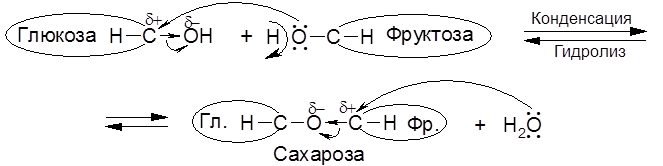
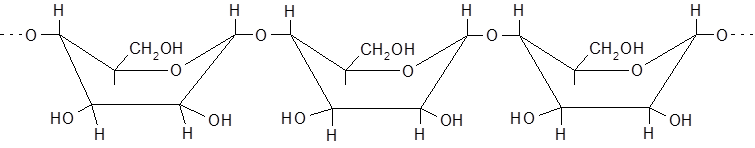 ,
,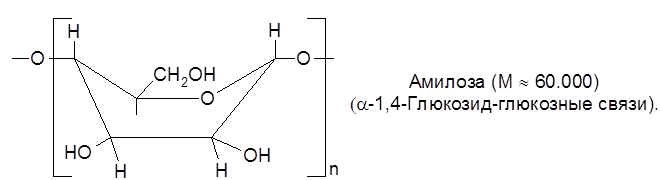
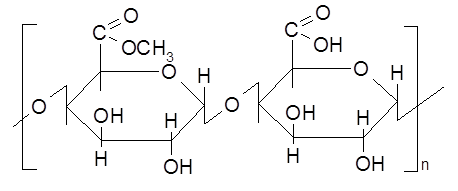

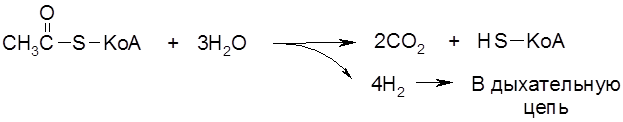
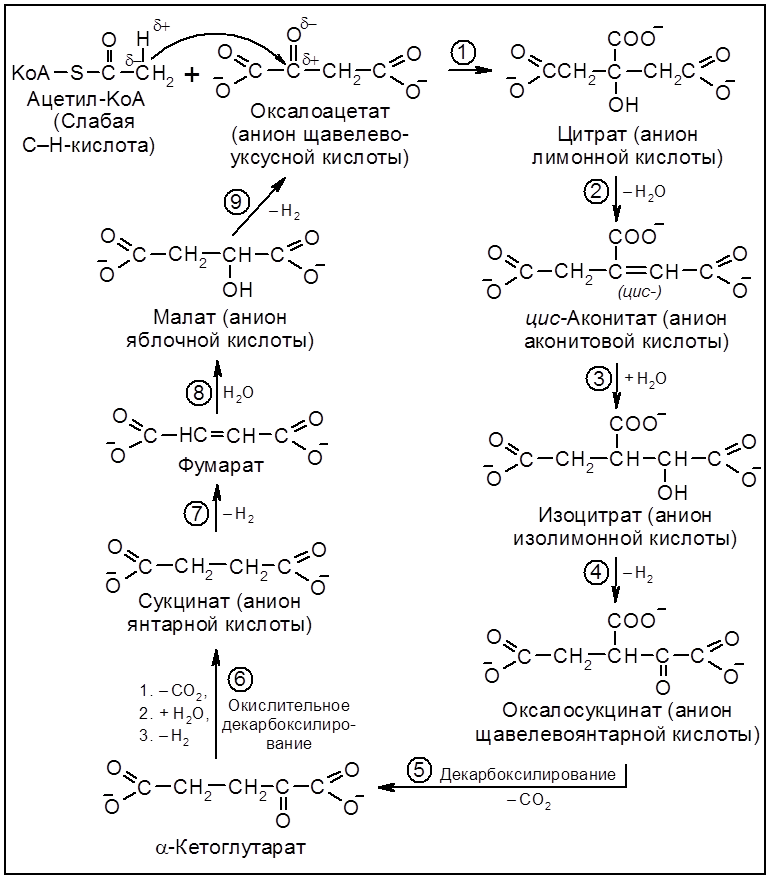
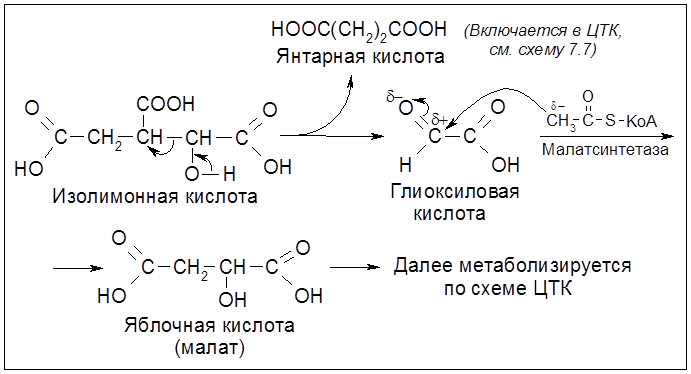
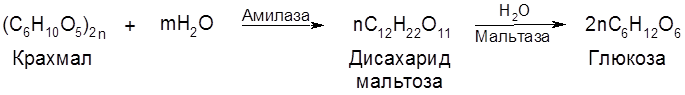
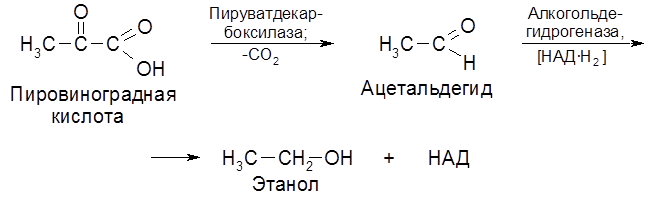
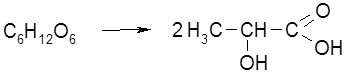
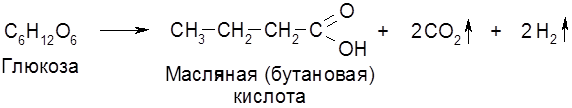
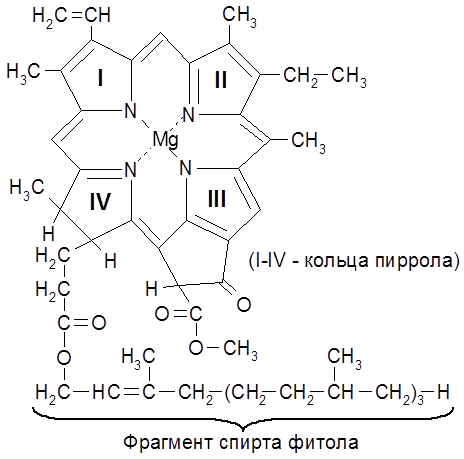
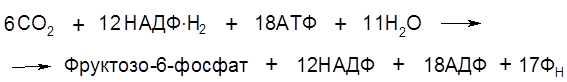
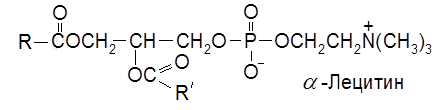
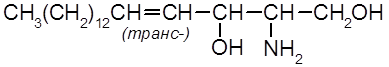
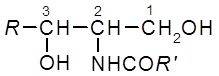
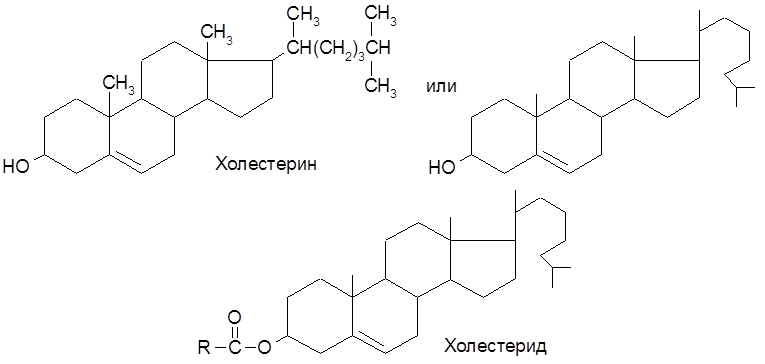
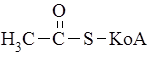 ) с СО2, в результате чего образуется производное малоновой кислоты – малонил-S-KoA:
) с СО2, в результате чего образуется производное малоновой кислоты – малонил-S-KoA:
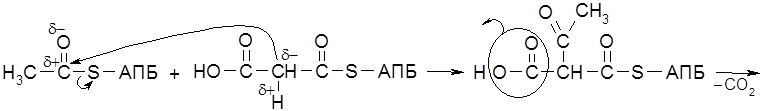

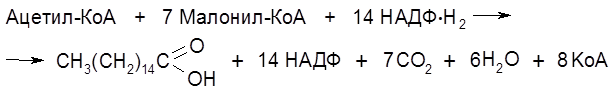
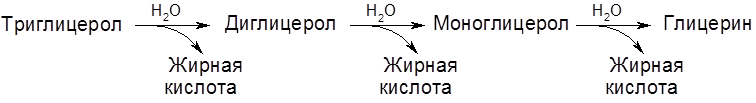
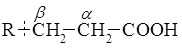 и последовательно отщепляют от нее двухуглеродные фрагменты в виде молекул ацетил-S-KoA (
и последовательно отщепляют от нее двухуглеродные фрагменты в виде молекул ацетил-S-KoA (